A befejezés a DNS replikáció utolsó fázisa. Ezt iniciáció és megnyúlás előzi meg. A replikáció korai leállítása rövidített fehérjék expresszióját és így mutációt eredményezhet.
Mi a felmondás?

A replikáció vagy redukció során a genetikai információhordozó DNS-t megsokszorozják az egyes sejtekben. A másolás a félig konzervatív elv szerint történik, és általában a genetikai információ pontos megismétléséhez vezet. A replikációt a szintézis fázisában, a mitózis fázis előtt indítják el, és így a sejtmag elosztása előtt zajlik.
A replikáció kezdetén a DNS kettős szálát egyszálra osztják, amelyeken új komplementer szálak alakulnak ki. Az egyes DNS-szálakat az ellenkező szál bázisszekvenciája határozza meg. A DNS replikációja több fázisban megy végbe. A megszűnés a replikáció harmadik és ezért utolsó fázisa. A megszüntetést iniciáció és megnyúlás előzi meg.
A megszűnés kifejezésének szinonim kifejezése ebben az összefüggésben a jelölés Felmondási szakasz. A megszűnés itt "megszakítást" vagy "megszüntetést" jelent. A lezárás során az újonnan kialakult mRNS-szál levál a tényleges DNS-től. A DNS-polimeráz munkája lassan véget ér. A DNS replikációjának befejezését nem szabad összekeverni az RNS replikációjának befejezésével.
Funkció és feladat
Az iniciáció replikációs szakaszában elsősorban a replikáció szabályozása zajlik. Meghatározzuk a replikáció kezdőpontját, és úgynevezett alapozásra kerül sor. Az inicializálás után megkezdődik a polimerizáció, amelynek során a meghosszabbítási fázist átvezetjük. Az enzim DNS-polimeráz elválasztja a DNS komplementer szálait egyetlen szálra, és egymás után leolvassa az egyes szálak alapjait. Ebben a fázisban félig szakaszos megduplázódás történik, amely magában foglalja az alapozás egy másik szakaszát is.
Csak a megindítás és meghosszabbítás után következik be a replikáción belül a terminációs szakasz. A megszűnés életformától kezdve eltérő. Az eukariótákban, mint például az emberek, a DNS gyűrűben van felépítve. Ide tartozik olyan terminációs szekvenciák is, amelyek két különböző szekvenciának felelnek meg, amelyek mindegyike releváns egy replikációs villához.
A megszüntetést általában nem indítják el speciális mechanizmusok. Amint két replikációs villa fut össze vagy a DNS véget ér, a replikáció ezen a ponton automatikusan véget ér. Így fejeződik be a replikáció automatikus mechanizmusban.
A lezárási szekvenciák vezérlő elemek. Biztosítják, hogy a replikációs szakasz ellenőrzött módon érkezzen meg egy adott végponthoz, annak ellenére, hogy a replikációs villák a különböző replikációs sebességeket mutatják. Minden terminációs hely megfelel a Tus fehérje, a "terminus hasznosító anyag" kötési helyeinek. Ez a protein blokkolja a DnaB replikációs helikázt, és ezzel megállítja a replikációt.
Az eukariótákban a replikált gyűrűs szálak a replikáció után is kapcsolódnak egymáshoz. A csatlakozás megfelel a végpontnak. Csak a sejtosztódás után különféle folyamatok választják el őket, és így megoszthatók. Úgy tűnik, hogy a sejtosztódásig megmaradó kapcsolat szerepet játszik a szabályozott eloszlásban.
Két fő mechanizmus játszik szerepet a DNS-gyűrűk végső elválasztásában. Az elválasztásban olyan enzimek, mint az I. és II. Típusú topoizomeráz vesznek részt. Végül egy segédfehérje felismeri a stop kodont a terminálás alatt. Ez azt eredményezi, hogy a polipeptid leesik a riboszómáról, mivel nem áll rendelkezésre t-RNS, amely a stop kodon számára megfelelő antikodont tartalmaz. Végül a riboszóma két alegységre bomlik.
Betegségek és betegségek
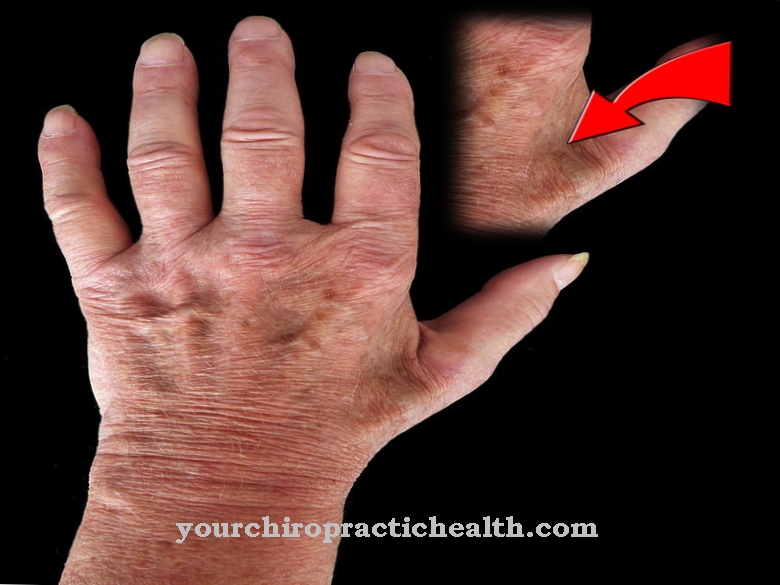
A replikáció szempontjából a genetikai anyag megduplázódásának minden folyamata bonyolult, és nagy mennyiségű anyagot és energiát igényel a sejtben. Ezért könnyen előfordulhatnak spontán replikációs hibák. Ha a genom spontán megváltozik, vagy kívülről indukálódik, akkor mutációkról beszélünk.
A replikációs hibák hiányzó bázisokat eredményezhetnek, megváltozott alapokkal társíthatók, vagy a helytelen bázispárosítás következményei lehetnek. Ezenkívül az egy vagy több nukleotid deléciója és beépítése a két DNS-szálba replikációs hibákhoz vezethet. Ugyanez vonatkozik a pirimidin dimerekre, a szál törésekre és a DNS-szálok térhálósítási hibáira.
Különböző javítási mechanizmusok érhetők el replikációs hiba esetén. A legtöbb említett hibát, amennyire csak lehetséges, kijavítják a DNS-polimeráz segítségével. A replikációs pontosság viszonylag magas. A hibaarány nukleotidonként csak egy hiba, amelyet a különböző kontroll rendszerek okoznak.
Például az eukarióta sejtek kontrollmechanizmusát nonszensz-közvetített mRNS-bomlásnak nevezzük, amely felismeri a nem kívánt stop kodonokat az mRNS-en belül, és így megakadályozza a rövidebb fehérjék expresszióját.
Az korai stop-kodonok az mRNS-ben a génmutációknak tudhatók be. Az úgynevezett nonszensz mutációk vagy az alternatív és hibás splicing rövidített fehérjéket eredményezhetnek, amelyeket funkcionális veszteségek érintnek. A vezérlő mechanizmusok nem mindig korrigálhatják a hibákat.
Az autoszomális recesszív öröklött rendellenesség három különféle formája létezik: β-talassemia: az első a homozigóta talassemia, egy súlyos betegség, amely visszavezethető a hülye mutációra. A heterozigóta talassemia enyhébb betegség, amelyben a nonszensz mutációk csak a β-globin gén egyetlen példányában találhatók meg. A nonszensz által közvetített mRNS-bomlás mechanizmusa révén a hibás gén mRNS-é olyan mértékben lebontható, hogy csak az egészséges gének expresszálódjanak.
Heterozigóta talassemia és így a betegség mérsékelt formájában a nonszensz mutáció az utolsó mRNS exonban van, így a kontroll mechanizmusok nem aktiválódnak. Ezért az egészséges β-globin mellett rövidített β-globin is kialakul.A hibás β-globin vörösvértestek elpusztulnak.
A kontrollmechanizmus kudarcának egy másik példája a Duchenne izomdisztrófia, amely szintén az mRNS hülye mutációjának következménye. Ebben az esetben a kontroll mechanizmus lebontja az mRNS-t, de ezáltal az úgynevezett dystrophin protein teljes veszteségét okozza.
.jpg)








.jpg)



.jpg)